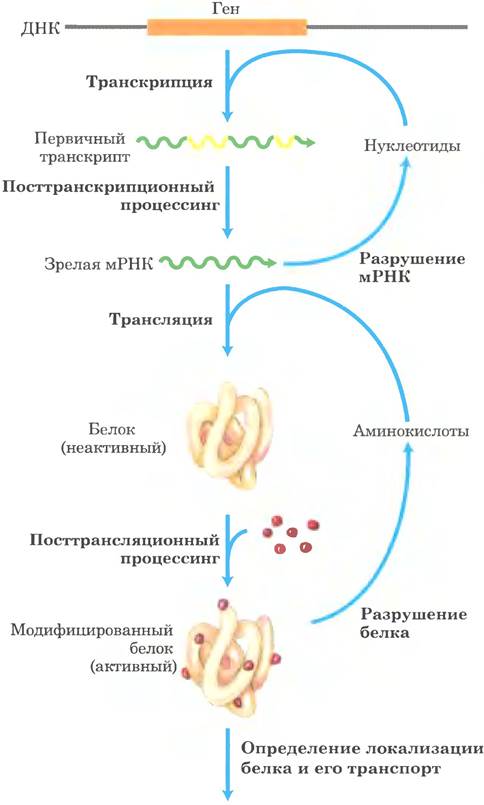
Транскрипция и трансляция[ | ]
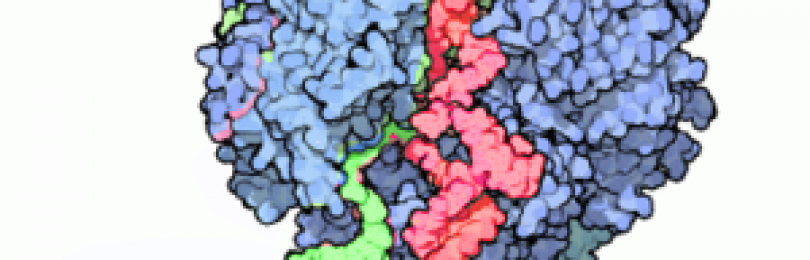
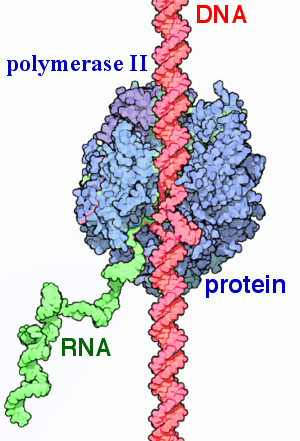 Транскрипция ДНК в РНК используя фермент РНК полимеразу II.
Транскрипция ДНК в РНК используя фермент РНК полимеразу II.
Основные статьи: Транскрипция (биология), трансляция (биология)
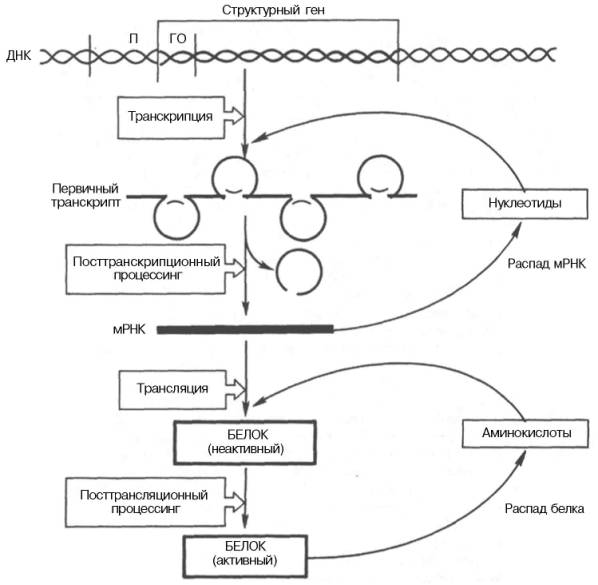
У прокариот и эукариот гены представляют собой последовательности нуклеотидов ДНК. На матрице ДНК происходит транскрипция — синтез комплементарной РНК. Далее на матрице мРНК происходит трансляция — синтезируются белки. Существуют гены, кодирующие нематричную РНК (например, рРНК, тРНК, малые РНК), которые экспрессируются (транскрибируются), но не транслируются в белки.
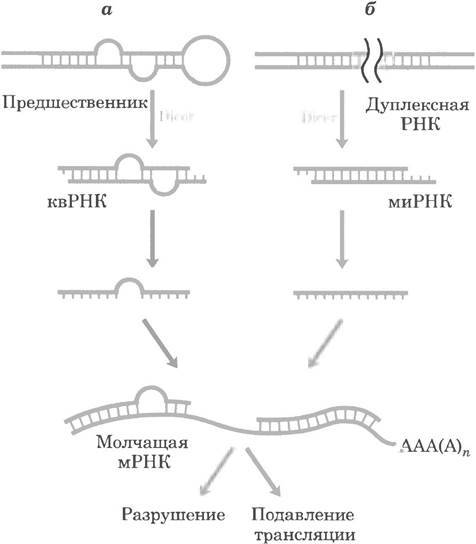
Регуляция после транскрипции |
Основная статья: МикроРНК
МикроРНК — это короткие (18—25 нуклеотидов) последовательности односпиральной РНК, вызывают подавление экспрессии генов. МикроРНК связываются со своей мишенью — информационной РНК — по принципу комплементарности. Это вызывает подавление синтеза белка или деградацию информационной РНК.
МикроРНК могут иметь большую или меньшую специфичность благодаря большей или меньшей доле комплементарных своей мишени азотистых оснований. Низкая специфичность позволяет одной микроРНК подавлять экспрессию сотен разных генов.
Репликация ДНК. Процесс репликации ДНК
Информация, записанная в ДНК, должна быть не только реализована в процессе развития клеток и организмов, но и в полном объеме передана следующему поколению. С этой целью перед делением клетки в ней осуществляется процесс репликации , т.е. удвоения количества ДНК.
Информация о механизме репликации содержится в самой ДНК: одни гены кодируют ферменты, синтезирующие предшественники ДНК — нуклеотиды, другие — ферменты, обеспечивающие соединение активированных нуклеотидов в единую цепочку. Механизм репликации был впервые постулирован Дж. Уотсоном и Ф. Криком, которые отмечали, что комплементарность цепей ДНК наводит на мысль, что эта молекула может удваивать саму себя. Они предположили, что для удвоения необходим разрыв водородных связей и расхождение цепей, каждая из которых играет роль матрицы при синтезе комплементарной цепи. В результате одного акта удвоения образуются две двунитиевые молекулы ДНК, в каждой из которых имеется одна материнская нить и одна новая (см. рис.).
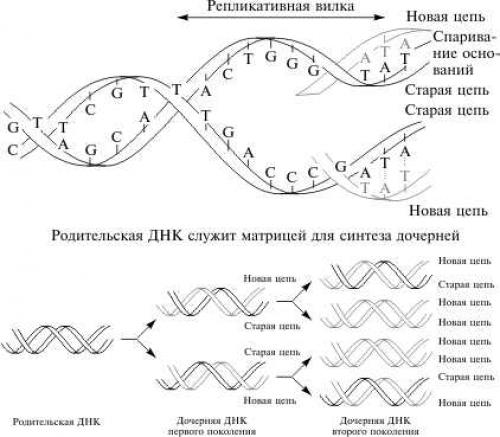
Полуконсервативная репликация ДНК
Механизм получил название полуконсервативной репликации . Позже матричная природа и постулированный принцип репликации ДНК были подтверждены многочисленными экспериментальными данными.
Репликация ДНК начинается в специфических точках хромосомы — сайтах инициации репликации (origin). Процесс репликации обслуживается большим количеством ферментов. Наиболее полно изучен аппарат репликации бактериальной ДНК, особенно E. coli. Функцию расплетания молекулы ДНК у прокариот выполняют специфические ферменты геликазы , которые используют для работы энергию гидролиза АТФ до АДФ. Они часто функционируют в составе белкового комплекса, осуществляющего перемещение вилки и репликацию расплетенных нитей. Удерживают нити ДНК от воссоединения другие специфические белки, связывающиеся с одноцепочечными участками. Эти участки, разошедшиеся в разные стороны, образуют характерную структуру — репликативную вилку (вилку Кернса). Это — та часть молекулы ДНК, в которой в данный момент осуществляется синтез новой цепи. В продвижении вилки большую роль играет белок гираза , относящийся к разряду топологических изомераз. Он обнаружен только у бактерий. Гираза — это релаксирующий фермент, который, производя двунитиевые разрывы, снимает положительные (перед вилкой) и способствует образованию отрицательных (сзади вилки) супервитков в релаксированной ДНК.
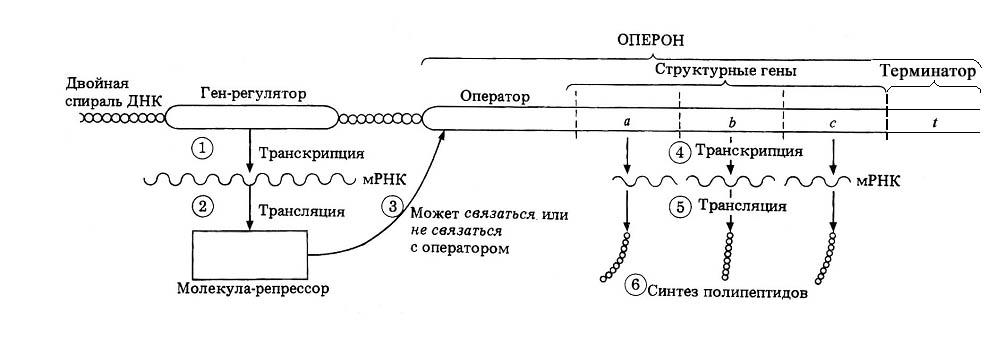
Оперон
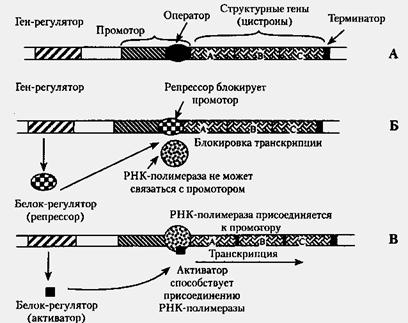
Оперон — функциональная единица генома у прокариот , в состав которой входят цистроны ( гены , единицы транскрипции), кодирующие совместно или последовательно работающие белки и объединенные под одним (или несколькими) промоторами . Такая функциональная организация позволяет эффективнее регулировать транскрипцию этих генов.
Опероны по количеству цистронов делят на моно-, олиго- и полицистронные, содержащие, соответственно, только один, несколько или много цистронов (генов).
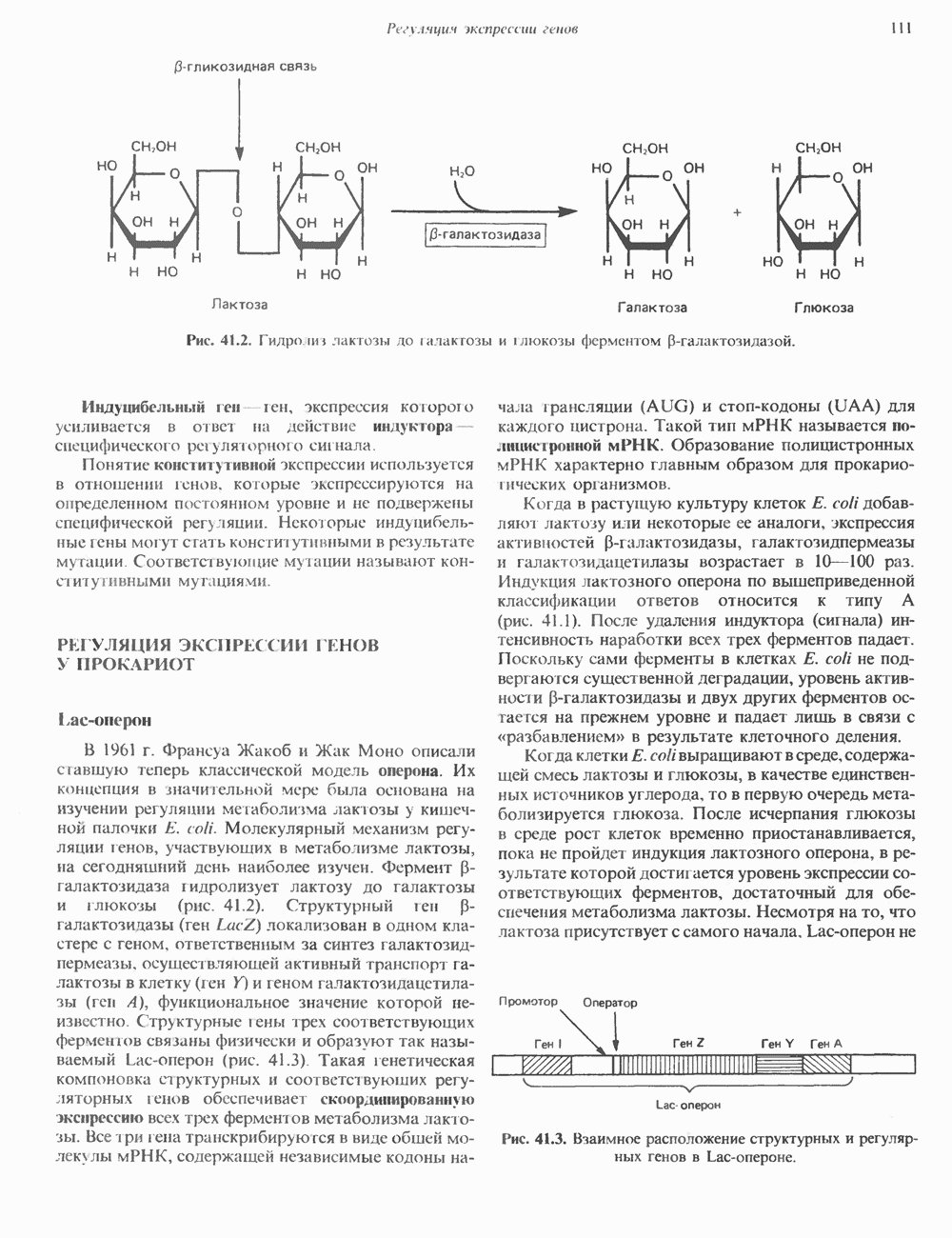
Характерным примером оперонной организации генома прокариот является лактозный оперон , триптофановый , пиримидиновый и bgl опероны у Escherichia coli
Начинается и заканчивается оперон регуляторными областями — промотором в начале и терминатором в конце, кроме этого, каждый отдельный цистрон может иметь в своей структуре собственный промотор и/или терминатор.
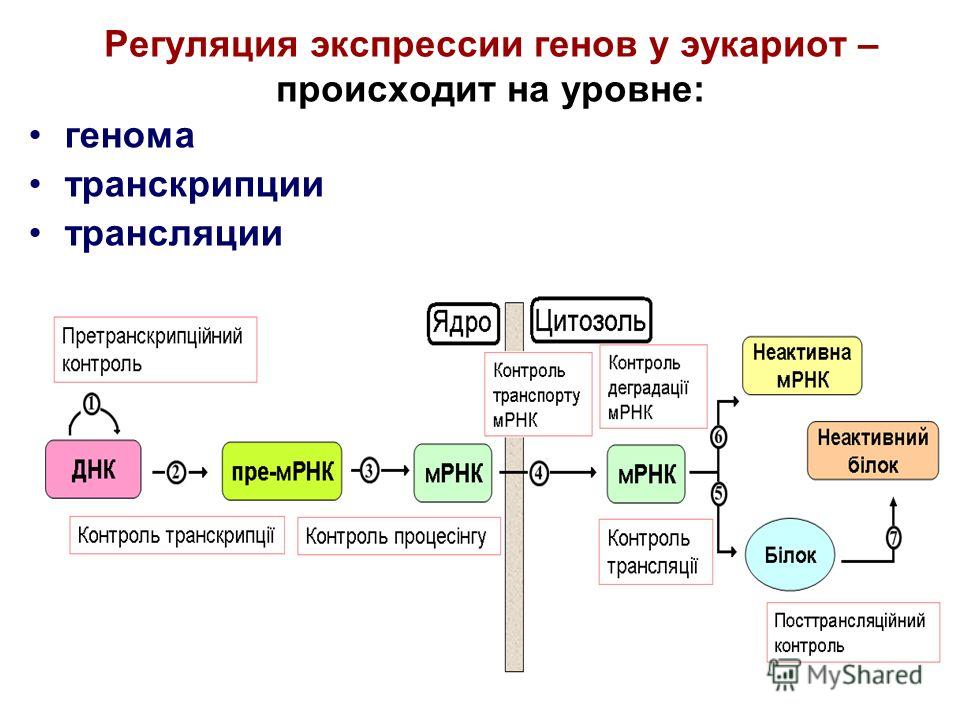
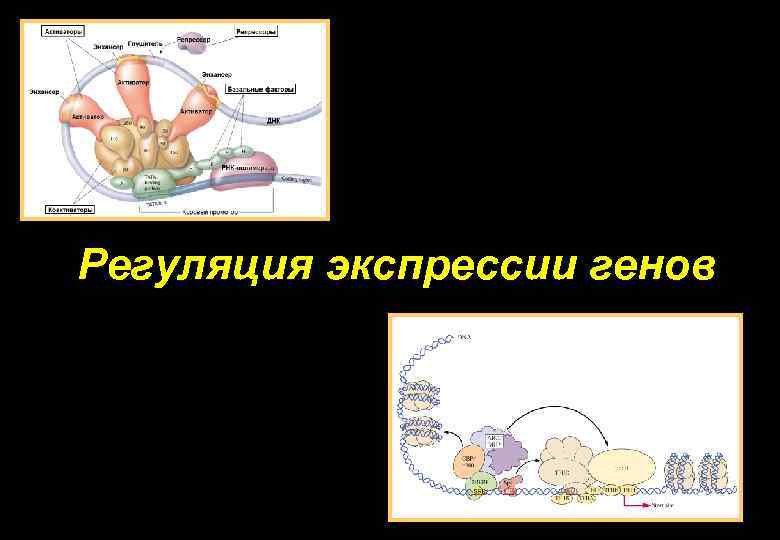
Регуляция экспрессии генов эукариот на уровне транскрипции. Регуляция экспрессии генов на уровне транскрипции у эукариот
Несмотря на то, что основные принципы регуляции транскрипции генов у прокариотических и эукариотических организмов остаются неизменными — через специфические взаимодействия белков и нуклеиновых кислот друг с другом, а также между собой, данный процесс у эукариот характеризуется рядом существенных особенностей. Это связано, прежде всего, с необходимостью поддержания координированной экспрессии эукариотических генов в более сложноорганизованной генетической системе. Достаточно вспомнить, что в организме человека гистологически различают, по крайней мере, 100 типов клеток, формирующих его органы и ткани. Для любого типа клеток характерен свой уникальный набор экспрессирующихся генов, которые начинают функционировать во время дифференцировки клеток-предшественников. Кроме того, сам процесс формирования органов и тканей сопровождается пролиферацией строго определенных групп клеток, а также упорядоченным во времени и пространстве перемещением клеток. Все эти особенности жизнедеятельности клеток высших организмов обеспечиваются функционированием их генов.
Регуляция экспрессии генов у эукариот
У многоклеточных организмов в клетках разных тканей экспрессируются разные гены, т.
е. для эукариот характерна дифференциальная экспрессия.

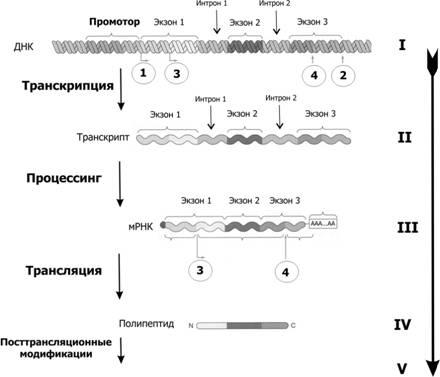
У эукариот, также как и у прокариот, существуют регуляторные белки с похожим механизмом действия. При этом для эукариот не характерна регуляция по типу оперона. Цистроны (транскрибируемые участки) эукариот обычно содержат по одному гену. (Это не касается геномов хлоропластов и митохондрий.)
Кроме регуляторных белков, взаимодействующих с ДНК, у эукариот существуют и другие способы регуляции экспрессии генов.
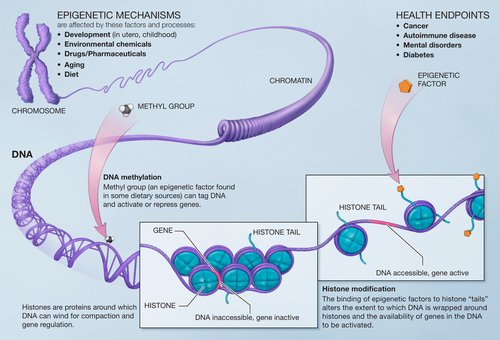
Конденсация и деконденсация хроматина. Это наиболее универсальный метод регуляции транскрипции.
Когда нужно экспрессировать определенные гены, хроматин в этом месте деконденсируется.
Альтернативные промоторы. У гена может быть несколько промоторов, каждый из которых начинает транскрипцию с разных его экзонов в зависимости от типа клетки. В конечном итоге будут синтезированы разные белки.
Метилирование и деметилирование ДНК. Метилирование ДНК происходит в регуляторных областях гена. Метилируется цитозин в последовательности ЦГ, после чего ген инактивируется.
При деметилировании активность гена восстанавливается. Процесс регулируется ферментом метилтрансферазой.
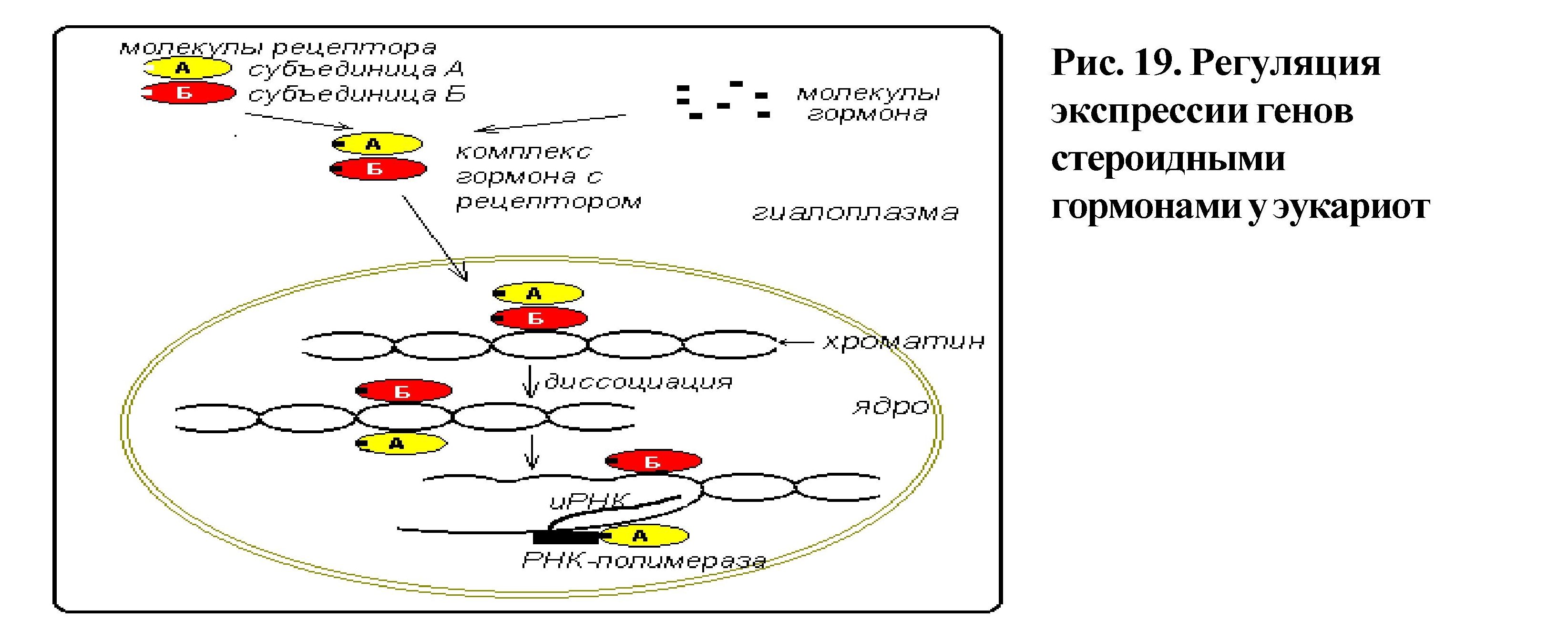
Гормональная регуляция. При гормональной регуляции гены активируются в ответ на внешний химический сигнал (поступление в клетку определенного гормона).
Этот гормон запускает те гены, которые имеют специфические последовательности нуклеотидов в регуляторных областях.
Геномный импринтинг. Это малоизученный способ регуляции экспрессии генов у эукариот. Он возможен только у диплоидных организмов и выражается в том, что активность генов зависит, от какого из родителей они были получены. Выключение генов осуществляется путем метилирования ДНК.
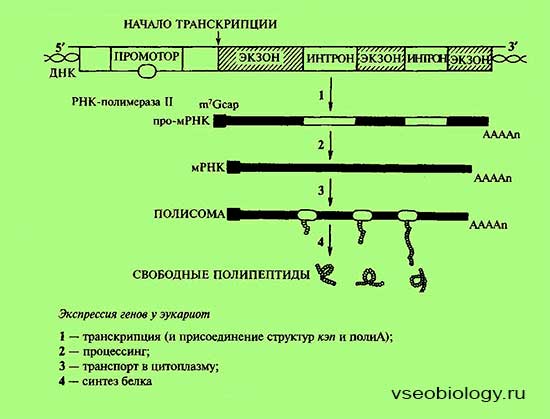
Альтернативный сплайсинг. Это регуляция на уровне процессинга.
При альтернативном сплайсинге порядок сшивки экзонов может быть различным. Отсюда следует, что на основе одной и той же нуклеотидной последовательности ДНК могут быть синтезированы разные белки. Хотя их отличие друг от друга будет в основном заключаться лишь в разных сочетаниях одних и тех же аминокислот.
Тканеспецифическое редактирование РНК также протекает на уровне процессинга. Выражается в замене отдельных нуклеотидов в РНК в определенных тканях организма.
Кроме того, у эукариот иРНК часто не подвергается процессингу вообще (а распадается) или подвергается с задержкой.
Это токже можно рассматривать как способ регуляции экспрессии генов.
Регуляция стабильности иРНК. У эукариот существует регуляция и на уровне трансляции, когда готовые иРНК не «допускаются» к рибосомам или разрушаются. Другие же иРНК могут дополнительно стабилизироваться для многократного использования.
Посттрансляционная модификация белка. Чтобы молекула полипептида превратилась в активную молекулу белка, в ней должны произойти различные модификации определенных аминокислот, должны быть сформированы вторичная, третичная и возможно четверичная структуры.




На этом этапе также можно повлиять на реализацию генетической информации, например, не дав молекуле сформироваться.
Риборегуляторы. Были обнаружены РНК, выполняющие регуляторные функции путем ослабления работы отдельных генов.
Для высокоорганизованных животных отмечается существование надклеточного уровня регуляции экспрессии генов.
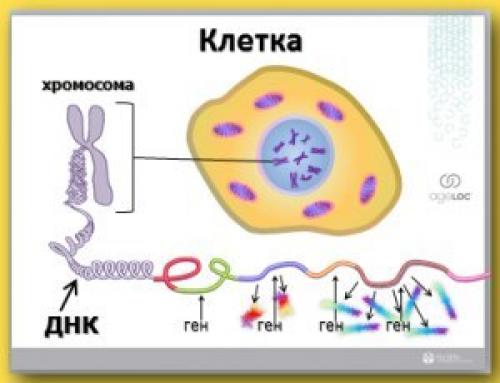
У этого термина существуют и другие значения, см. Экспрессия.
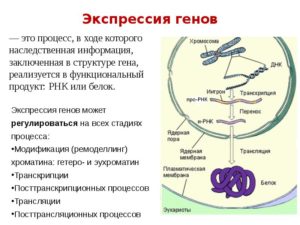
Экспрессия генов — это процесс, в ходе которого наследственная информация от гена (последовательности нуклеотидовДНК) преобразуется в функциональный продукт — РНК или белок.
Некоторые этапы экспрессии генов могут регулироваться: это транскрипция, трансляция, сплайсинг РНК и стадия посттрансляционных модификаций белков. Процесс активации экспрессии генов короткими двуцепочечными РНК называется активацией РНК.
Регуляция экспрессии генов позволяет клеткам контролировать собственную структуру и функцию и является основой дифференцировки клеток, морфогенеза и адаптации.
Экспрессия генов является субстратом для эволюционных изменений, так как контроль над временем, местом и количественными характеристиками экспрессии одного гена может иметь влияние на функции других генов в целом организме.
Метилирование ДНК. Вступление. Открытия лаборатории Белозерского.
В конце 1960-х годов в МГУ, на одном из семинаров межфакультетской лаборатории биоорганической химии, сотрудник А.Н. Белозерского , Борис Ванюшин, занимавшийся изучением необычных нуклеотидов в составе ДНК, высказал идею о том, что некоторые из этих необычных нуклеотидов, а именно 5-метилцитозин, играют важную роль в механизмах регуляции экспрессии генов, клеточной дифференцировки и процессов старения организма.
Образование этого 5-метилцитозина в ДНК было обусловлено ферментативным метилированием обычного цитозина, то есть посадкой на него метильных групп. ДНК в месте посадки на её цитозин метильных групп несколько изменяла свои свойства.
Количество 5-метилцитозина в ДНК менялось в разные периоды жизни организма от максимального, при рождении, до минимального, у дряхлого от старости. Так впервые в СССР была сформирована идея о том, что старение на уровне генома регулируется эпигенетически ( ) – то есть путём механизма вызывающего длительное и стабильное изменение экспрессии генов, которое не связано с изменениями в последовательности ДНК. В последующем Ванюшин опубликовал цикл работ под названием «Метилирование ДНК – эпигенетический контроль за генетическими функциями организма» и в 1977 году вместе с Г. Бердышевым книгу “Молекулярно-генетические механизмы старения”.
К сожалению, из-за сложности и затратности, используемой в те годы технологии изучения процессов метилирования ДНК, дальнейшие исследования в этом направлении были резко сужены.
Сегодня имеется множество разных методов определения метилирования ДНК, некоторые из которых (прохождение цепи ДНК через микропору генноинженерного белка порина А) позволяют читать результаты, не разрушая при этом ДНК, что намного упростило и удешевило подобные исследования ()
Экспрессия гена
Например, техника флюоресценцентной in situ гибридизации (FISH) последовательно демонстрировала зависимость между амплификацией HER-2/neu гена и рецидивом рака молочной железы и связанной с заболеванием смертью как в локализованных, так и в метастатических случаях (Рисунки 1 и 2).
ELISA измерения HER-2/neu протеина в клетках рака молочной железы также коррелируют с исходом заболевания. HER-2/neu в сыворотке может также оцениваться методом ELISA, хотя согласие относительно клинической полезности этой техники не достигнуто.
Методы southern- и slot-блоттинга менее эффективны, поскольку ДНК опухолевых клеток, экстрагированная из первичного образца карциномы, разбавляется ДНК из доброкачественной ткани молочных желез и воспалительных клеток. Последние исследования поддерживают ассоциацию между амплификацией HER-2/neu гена и гиперэкспрессией протеина и плохим клиническим исходом у пациентов с опухолями ЖКТ, легких и мочеполовой системы.
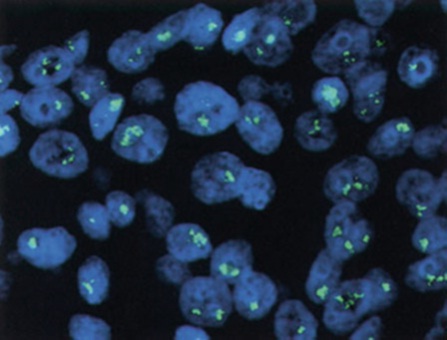
Фигура 1. Амплификация HER-2/neu гена, определяемая FISH.








Видны группы сигналов в каждом клеточном ядре
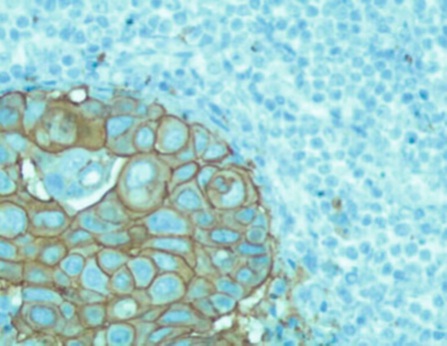
Фигура 2. Гиперэкспрессия HER-2/neu протеина, определяемая иммуногистохимией. Видно интенсивное окрашивание мембран
Таблица 1. Реактивы для HER-neu анализа
|
Название продукта |
Источник |
Метод |
Показание |
|
Прогноз |
|||
|
Ответ на герцептин |
|||
|
Прогноз, ответ на таксотер |
|||
|
Ответ на герцептин |
|||
|
Серологический тест |
Bayer Diagnostics |
Прогноз, ответ на терапию |
Клиническая эффективность препарата трастузумаба (Герцептина) стимулировала HER-2/neu тестирование в лечении рака молочной железы.
Трастузумаб, гуманизированное моноклональное антитело, вводится внутривенно и особенно эффективен в комбинации с цитотоксическими агентами (либо таксотер, либо Адриамицин+Цитоксан).
27%-ая частота отклика с этими режимами наблюдается у пациентов с прогрессирующим метастатическим раком молочной железы, рефрактерным к традиционному лечению. В 1998 FDA одобрил использование трастузумаба для лечения метастатического HER-2/neu-положительного рака молочной железы (IHC 2+ или 3+).







В 2006 FDA также одобрил трастузумаб в комбинации с цитотоксической химиотерапией для адьювантного лечения первичного рака молочной железы с метастазами в лимфатические узлы. Продолжающиеся клинические исследования трастузумаб+цитотоксическая терапия пока не показали достоверных результатов у пациентов с раком простаты, легкого, яичника и поджелудочной железы.
В дополнение к его роли в предсказании прогноза и ответа на терапию трастузумабом, HER-2/neu тестирование при раке молочной железы используется для выбора других вариантов лечения, включая использование HER-1/HER-2 ингибитора, лапатиниба.
Хотя способность HER-2/neu статуса предсказывать ответ на антиэстрогенную терапию не достигла согласия, повышенный ответ HER-2/neu-положительных опухолей на химиотерапевтические режимы, включающие антрациклины, обычно широко используется в клинической онкологии.
Однако, кардиотоксичность — основной побочный эффект трастузумаба — ограничивает его использование в комбинации с антрациклинами, особенно при метастатической форме заболевания.
Сложная экспрессия генов[ | ]
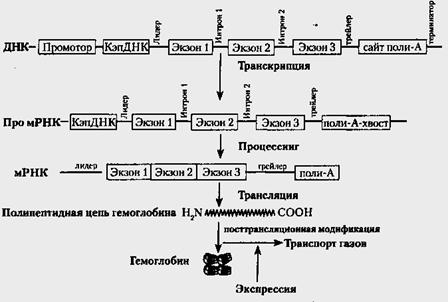
Примером сложной экспрессии генов в онтогенезе может служить генный контроль синтеза гемоглобинов у человека. Молекула гемоглобина состоит из 4 частей: двух идентичных альфа-цепей и двух идентичных бета-цепей. Гемоглобин нормального взрослого человека (НвА) отличается от гемоглобина эмбриона человека (эмбриональный гемоглобин, НвF). Различия между ними касаются бета-цепи. В гемоглобине плода она заменена на полипептидную гамма-цепь. Наконец, в крови взрослых людей в небольшом количестве встречается НвА2, в котором бета-цепь заменена на сигма-цепь. Все 3 типа нормальных гемоглобинов человека(НвА НвА2 НвF) контролируются отдельными локусами. Локус αА определяет формирование альфа-цепей. Он эффективен в течение всей жизни, обеспечивая наличие альфа-цепей во всех указанных гемоглобинах.
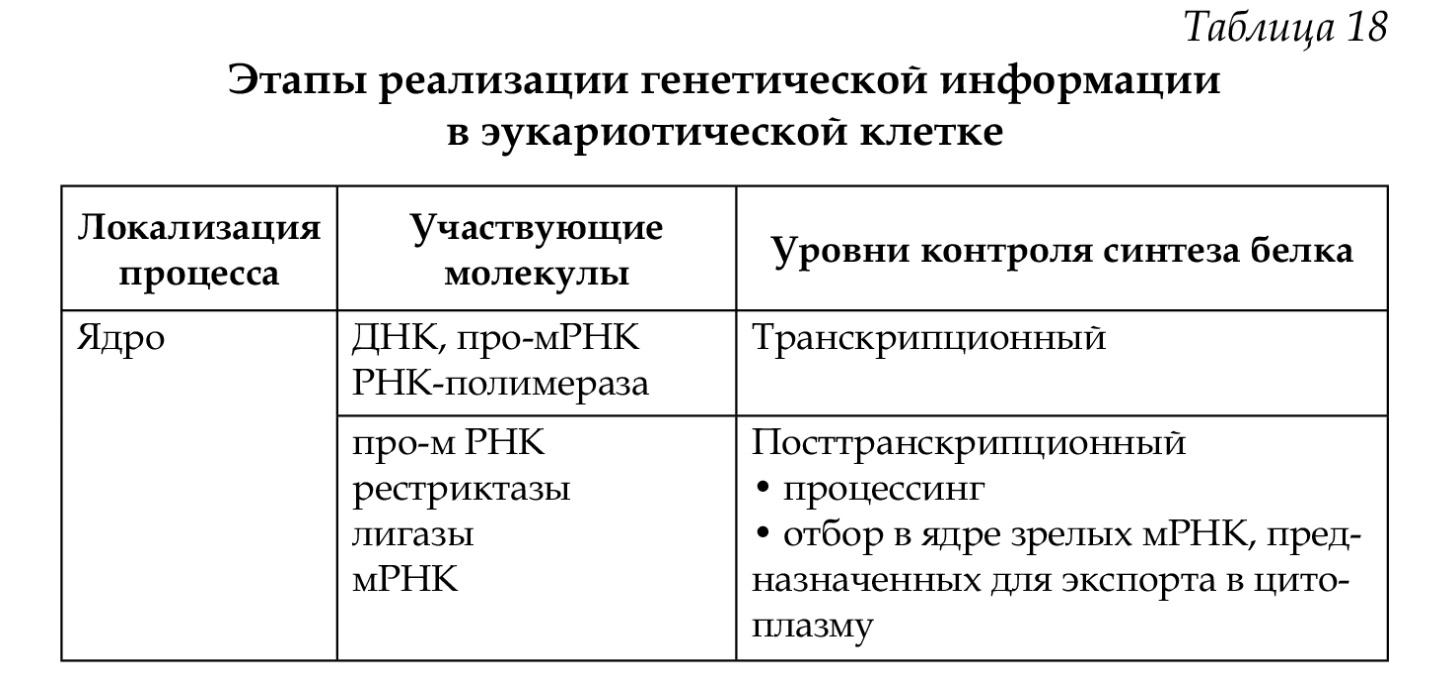
Общие принципы генетического контроля экспрессии генов
Важнейшим фактором регуляции генной активности являются элементы генома, отвечающие за синтез регуляторных
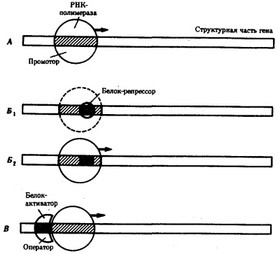
Рис. 3.84. Схема конститутивных и регулируемых генов.
А — конститутивный ген; Б, В — регулируемые гены; Б,, Б2— связывание РНК-полимеразы с промотором возможно лишь в отсутствие белка-репрессора, который специфически соединяется с оператором, частично или полностью перекрывающим промоторную последовательность; В — связывание РНК-полимеразы с промотором облегчается белком-активатором (апоиндуктором), который специфически узнает область оператора, расположенную перед промотором; стрелкой обозначено направление транскрипции
белков, — гены-регуляторы. Соединяясь с определенными нуклеотидными последовательностями ДНК, предшествующими структурной части регулируемого гена — операторами, белки-регуляторы способствуют или препятствуют соединению РНК-полимеразы с промотором. Если белок-регулятор взаимодействует с оператором, занимающим часть промотора или расположенным между ним и структурной частью гена, то это не дает возможности РНК-полимеразе соединиться с промоторной последовательностью и осуществить транскрипцию. Такой белок называют репрессором, и в этом случае осуществляется негативный контроль экспрессии гена со стороны гена-регулятора (рис. 3.85). Если промотор обладает слабой способностью соединяться с РНК-полимеразой, а ему предшествует область, узнаваемая белком-регулятором, присоединение последнего непосредственно перед промотором к молекуле ДНК облегчает связывание РНК-полимеразы с промотором, вслед за чем следует транскрипция. Такие белки называют активаторами (или апоиндукторами) , а контроль экспрессии гена со стороны гена-регулятора — позитивным (см. рис. 3.85).
Моноаллельная экспрессия генов
Моноаллельная экспрессия у эукариот характерна:
- для генов Х-хромосомы в женских клетках из-за механизма дозовой компенсации;
- для импринтируемых генов;
- В настоящее время известно, что около 5—10 % генов эукариот экспрессируются в клетках моноаллельно, среди таких генов чаще наблюдаются гены, кодирующие поверхностные клеточные белки и, в частности, гены, кодирующие иммуноглобулины, Т-клеточные и обонятельные рецепторы.
Это явление носит также название аллельное исключение.
Выбор экспрессирующегося аллеля происходит рано в развитии, и этот выбор осуществляется случайно, в результате около половины клеток организма экспрессируют отцовский аллель, а другая половина клеток — материнский аллель. Иногда наблюдается тканеспецифичная моноаллельная экспрессия гена, в других тканях такой ген может экспрессироваться биаллельно. К случайной моноаллельной экспрессии аутосомных генов не относят случаи, когда разные аллели гена экспрессируются на различном уровне из-за полиморфизма в cis-регуляторных последовательностях гена.
Литература
- Патрушев Л. И. Экспрессия генов. — М.: Наука, 2000. — ISBN 5-02-001890-2
- Бекиш О.-Я. Л. Медицинская биология. — Минск: Ураджай, 2000. — 518 с.
CC wikiredia.ru
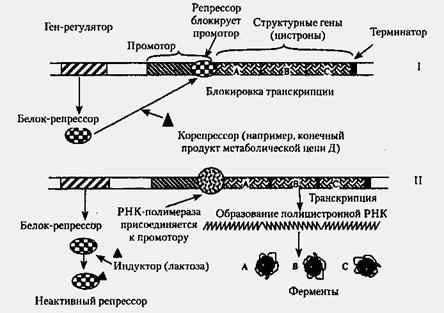
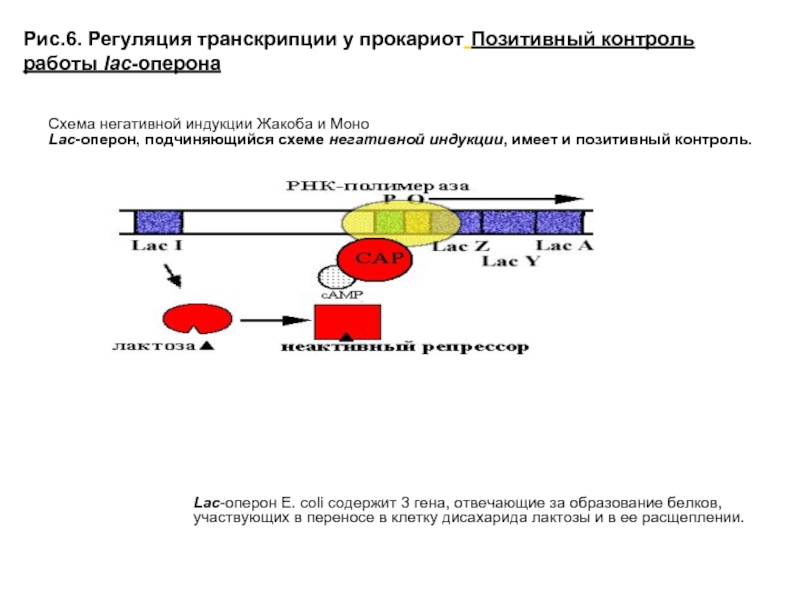
Позитивная и негативная Регуляция экспрессии генов. Регуляция экспрессии генов у прокариот
У прокариот пока молекула РНК синтезируется на участке ДНК, она тут же может транслироваться (начиная с уже синтезированного конца). Поэтому у них регуляция экспрессии (активности) генов осуществляется почти исключительно на уровне ДНК, так как в РНК часто невозможно внести какие-нибудь изменения до ее трансляции.
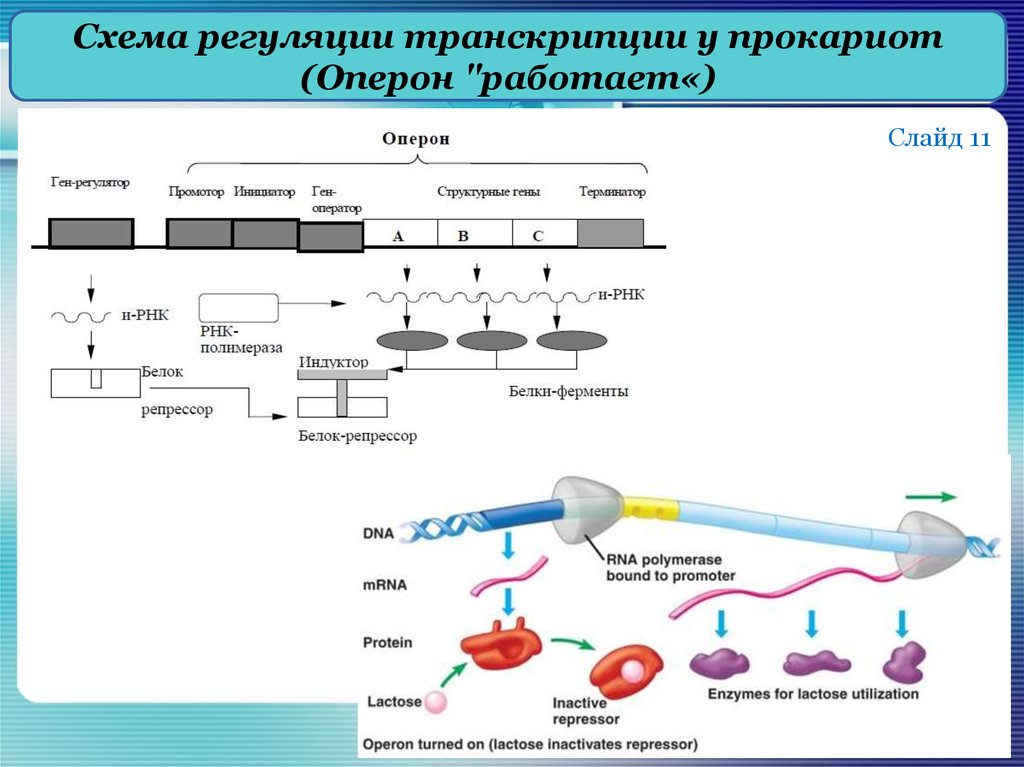
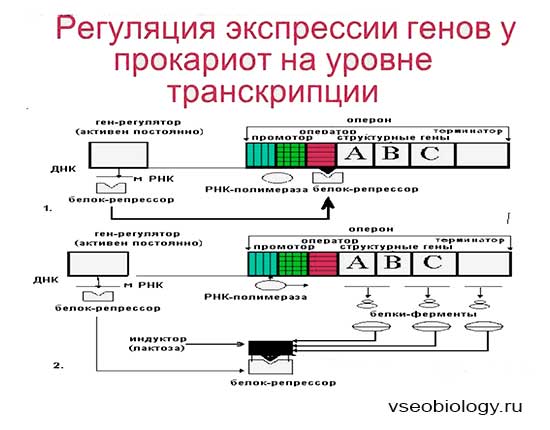
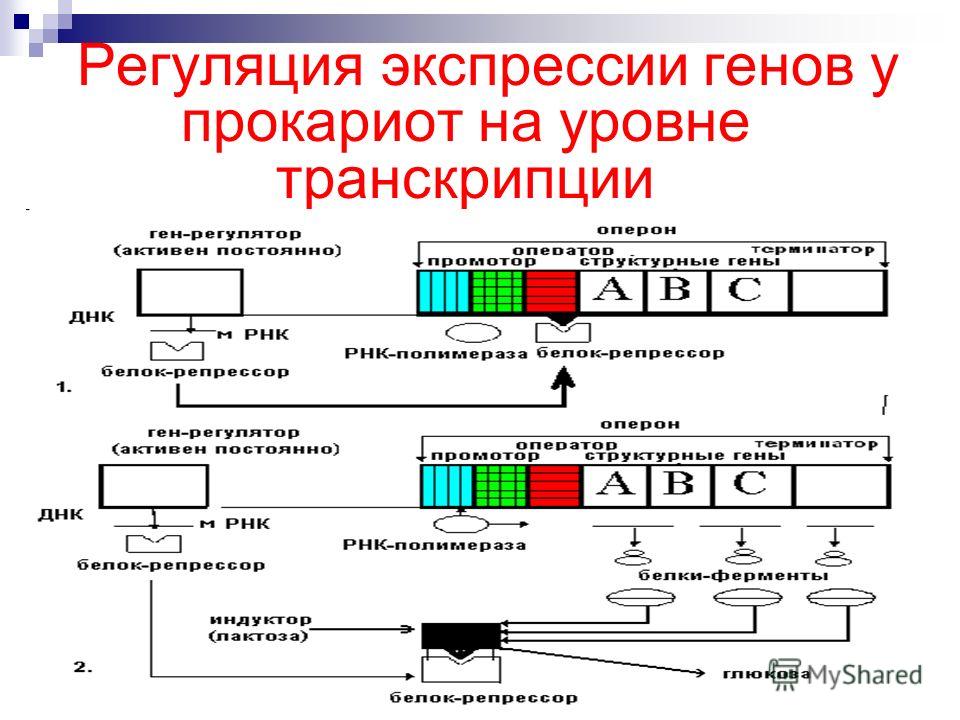
В 1961 г. Жакобом и Моно была предложена модель оперона как системы регуляции генов у бактерий. Оперон состоит из промотора, оператора, структурных генов оперона (их может быть разное количество) и терминатора. В области промотора прикрепляется фермент РНК-полимераза. В области оператора присоединяется белок-репрессор, который кодируется отдельно отстоящим от оперона геном-регулятором (может быть сцеплен со своим опероном, а может находиться на расстоянии).
Если белок-репрессор соединяется с оператором, то транскрипция всех структурных генов оперона становится невозможной, так как РНК-полимераза не может перемещаться по цепи ДНК.
В свою очередь активность белка-репрессора может блокироваться определенным для него низкомолекулярным соединением — индуктором (тем или иным питательным веществом бактерий). В результате взаимодействия с индуктором белок-репрессор видоизменяется и уже не может присоединиться к оператору своего оперона. В этом случае гены оперона экспрессируются (т. е. на них идет синтез).




Бывает обратная ситуация, когда индуктор активирует белок-репрессор.
Таким образом, в зависимости от того, какие индукторы находятся в цитоплазме, у прокариот экспрессируются те или иные генные группы.
Вышеописанный механизм экспрессии генов относится к негативной регуляции, так как гены транскрибируются, если они не выключены репрессором. И наоборот: не транскрибируются, если выключены.
Кроме негативной регуляции у бактерий существует также позитивная. В этом случае вместо белка-репрессора действие оказывает белок-активатор. На эти белки также действуют индукторы, активируя или инактивируя их.
Также у прокариот были выявлены опероны, которые актируются двумя регуляторными белками, соединенными друг с другом.
Экспрессия в медицине. Определение экспрессии
Что такое экспрессия? В медицине это генетический процесс, посредством которого содержащаяся в генах информация преобразуется в определенные клеточные структуры. Экспрессивность в биологии – это неразрывно связанное с жизнедеятельностью и здоровьем человека определение, которое получает дальнейшее развитие в психологии и психотерапии. К этой же медицинско-биологической сфере относится понятие «экспрессировать» – это означает транскрибировать и транслировать ген.
Экспрессивность – это степень выраженности определенного состояния, чувства, эмоции, отношения. Понятия «экспрессивность» и «экспрессия» применяются не только в психологии, но и в театроведении, искусствоведении, если требуется подчеркивать уровень выраженности духовности. А также показать средства выражения, к примеру, живопись, музыка, архитектурные элементы. Получается, что в имеющихся определениях данных терминов есть указания на связь этого явления с душевным и духовным миром людей.
Противоположным по смыслу значением является импрессивность – склонность к внутренним, а не внешним переживаниям, накоплению преимущественно отрицательных чувств. О таких людях говорят, что они держат переживания в себе, не склонны делиться эмоциями и проблемами, обиды и расстройства к ним приходят не сразу, не моментально.
Мнения о связи внутреннего мира людей и экспрессии, которые сформировались по большей части в искусствоведении и философии, дополняются психологией. Сущность этой связи в психологическом выражении видна в том, что экспрессия получает место не только сопровождения действий внешне. Она выступает частью данных явлений, формой их проявления.

Экспрессивная девушка рассказывает что-то своей подруге
По этой причине стоит говорить об экспрессии в виде личностного образования , способа познания духовного мира личности, а также в качестве внешнего «Я».